
Анатомия беспозвоночных пиявка прудовик дрозофила таракан рак
Серия: "Учебники для вузов. Специальная литература"
Издательство: "Лань" (1999)
Формат: 60x90/16, 320 стр.
Николай Фомичев
Орден Преподобного Сергия Радонежского I степени
Архиепи́скоп Ни́кон (в миру — Николай Васильевич Фомичёв, 9 мая 1910, Санкт-Петербург — 13 апреля 1995, там же) — деятель Русской православной церкви, архиепископ.
Содержание
Биография
Посещал братство при Александро-Невской Лавре, которым руководил архимандрит Варлаам (Сацердотский). Члены этой общины активно участвовали в жизни Церкви, пели на клиросе, организовывали чтение лекций и прочее. Участие в деятельности братства сыграло большую роль в формировании мировоззрения Николая Васильевича, а самое главное — в укреплении его веры.
В 1928 году окончил 102-ю трудовую школу, по окончании которой в течение трёх лет работал на производстве.
В 1931 году поступил в Ленинградский институт инженеров железнодорожного транспорта, который окончил в 1936 году с отличием, получив звание инженера-энергетика.
17 июня 1945 года митрополитом Ленинградским и Новгородским Григорием рукоположен во диакона, а 12 мая 1946 года — во священника. Священствовал в храмах г. Ленинграда.
В 1946 году поступил в Ленинградскую духовную академию, которую окончил экстерном в 1950 году со степенью кандидата богословия и назначен настоятелем Князь-Владимирского храма посёлка Лисий Нос. С 1952 года — настоятель Крестовоздвиженского собора и благочинный г. Петрозаводска Олонецкой епархии.
В 1953 году переехал служить на Украину и был назначен настоятелем Спасо-Преображенского собора в г. Житомире и секретарём епископа.
С 1956 года — настоятель кафедрального собора г. Великие Луки, секретарь епископа и благочинный. В 1958 году назначен настоятелем храмов в г. Новая Ладога Ленинградской епархии, а с конца 1960 года — в храмах Ленинграда на Большой Охте. С 1 февраля 1962 года был настоятелем Свято-Троицкого собора Александро-Невской Лавры.
11 августа 1962 года в Свято-Троицкой Лавре пострижен в монашество, 14 августа того же года возведён в сан архимандрита. 26 августа 1962 года хиротонисан во епископа Выборгского, викария Ленинградской епархии. С 16 ноября 1962 года — епископ Лужский, викарий Ленинградской епархии.
С 3 августа 1963 года — епископ Рижский и Латвийский. Вступил в конфликт с властями в связи с тем, что не дал своего согласия на закрытие Троице-Сергиева женского монастыря в Риге.
С 11 июня 1977 года — епископ Калужский и Боровский. 2 сентября 1977 года возведён в сан архиепископа. В этот период давление на церковь не прекращалось: старостой кафедрального собора были запрещены крестины, а на Пасху 1981 года Крестный ход проходил в сопровождении множества милицейских собак.
16 июля 1982 года назначен архиепископом Пермским и Соликамским. 28 марта 1984 года из-за постоянных конфликтов с местным уполномоченным совета по делам РПЦ был уволен на покой.
С 1984 года жил в Ленинграде (Санкт-Петербурге). С назначением на Ленинградскую кафедру митрополита Алексия (Ридигера) получил возможность совершать богослужения в храмах города и области, принимать участие в епископских хиротониях, рукополагать священнослужителей. Проповеди его отличались простотой и доступностью.
Награды
- Орден Преподобного Сергия Радонежского I степени (25 августа1982)
Библиография
- Биография
- Господь хранил верного раба Своего… Часть 1.
- Господь хранил верного раба Своего… Часть 2.
Евгений Поляков
Евгений Валентинович Поляков
Биография
- 1977—1986 — научный сотрудник, Уральский политехнический институт им. С. М. Кирова, Физико-технический факультет, кафедра радиохимии, Свердловск;
- 1986—1987 - доцент, Уральский политехнический институт им. С. М. Кирова, Физико-технический факультет, кафедра радиохимии, Свердловск;
- 1988—1994 - старший научный сотрудник лаборатории физико-химических методов анализа, институт химии твердого тела УрО РАН, Екатеринбург;
- С 1994 — заведующий лабораторией лаборатории физико-химических методов анализа, институт химии твердого тела УрО РАН, Екатеринбург;
- C 2004 — заместитель директора (по научным вопросам), институт химии твердого тела УрО РАН, Екатеринбург.
1. Drosophila melanogaster, или плодовая (уксусная) муха
Дрозофила – маленькое насекомое семейства Drosophiliadae из отряда Diptera. Наблюдать этих симпатичных мушек можно около испорченных фруктовых отходов – вокруг этого растительного материала развивается своя особая жизнь. Они настолько малы (1,5-4 мм), что подчас их можно и не заметить.
Однако это крохотное существо вошло в историю науки: уже более ста лет дрозофила используется как модельный объект для различных исследований области генетики и, в последнее время, биологии развития. Ее важность для здоровья человека была отмечена Нобелевской премией, врученной медикам-физиологам Ed Lewis, Christiane Nusslein-Volhard и Eric Wieschaus в 1995 году.
В мире существует более 1000 видов этих прелестных существ, причем наиболее распространены они в субтропиках и тропиках. На Гавайских островах, например, мушиное население включает свыше 300 видов, а вот на территории нашей страны их на порядок меньше. Родиной этих мушек считается Индо-Малайская область. В настоящее время этот вид широко распространен, в том числе на Кавказе и Украине.
2. Морфология и половой деморфизм
Дикий тип дрозофилы имеет ярко-красные глаза и “серое” тело. Нормально развитые крылья превышают длину тела. Питаются в природе мушки соком растений, гниющими растительными остатками, но в лаборатории их разводят на специально приготовленной среде.
Самки и самцы морфологически отличаются друг от друга по ряду признаков. Самки несколько крупнее самцов, однако, размеры мушки во многом зависят от условий питания, особенно на личиночной стадии. Брюшко у самки немного округлое с заостренным концом, у самца оно цилиндрическое с притупленным концом. Последние несколько сегментов у самца сплошь пигментированы, в отличие от самки. К тому же, у самки имеется восемь хорошо развитых верхних хитиновых щетинок груди, называемых тергитами, а у самца – шесть: шестой и седьмой срослись, а восьмой вошел в состав полового аппарата. Стернитов, то есть хитиновых пластинок с брюшной стороны, у самки насчитывается четыре. А у самца – три, причем у представителей каждого пола первый, второй, седьмой и восьмой недоразвиты. Хитиновые щитки тергиты, стерниты и плейриты участвуют в движении крыльев.
К числу вторичных половых признаков у самца относятся половые гребешки, представляющие собой крепкие хитиновые щетинки на первом членике лапки передних ног. У самки подобные образования отсутствуют.
3. Биология развития
Яйца дрозофилы несколько вытянуты (около 0,5 мм в длину) и хорошо заметны, будучи отложенными на участках питательной среды с пониженной влажностью. Яйцо защищено двумя оболочками, различными по происхождению. Внутренняя, образованная самим яйцом, называется желточной, а наружная – хорион – продукт фолликулярного эпителия яичника. У яйца дрозофилы различают дорсальную и вентральную стороны, задний и передний конец. От передней части дорсальной стороны отходят и идут по направлению кпереди два жгутикообразных выроста хориона, предохраняющих яйцо от попадания в жидкую среду.
В нормальных условиях эмбриональное развитие протекает вне тела матери при температуре 27 0 около 20 часов. В благоприятных условиях каждая самка откладывает до 50-80 яиц в сутки, а всего в течение 3-4 суток она может отложить их более 200.
Личинка после вылупления из яйца начинает усиленно питаться микроорганизмами, избыток пищи определяет размеры и жизнеспособность будущего организма. После недолгого

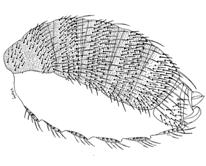
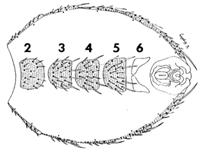
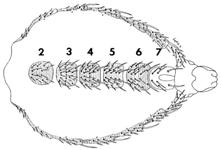
 |
Личинки некоторое время оживленно ползают по стенкам пробирки, ничем не питаясь, - начинается стадия окукливания. Затем они становятся неподвижными, значительно уменьшаются в длину и приобретают бочкообразную форму. Личиночные органы и ткани, за исключением гонад и нервной системы, разрушаются, а органы взрослой мухи начинают развиваться из имагинальных дисков.
По окончании третьих суток становятся заметны очертания глаз с желтоватым пигментом. Вылупление мухи и разрыв куколки достигается нагнетанием жидкости. Молодые, с длинным желтоватым телом, без пигмента и короткими, еще не расправленными крыльями, мухи обычно выходят из куколки рано утром.
4. Дрозофила как научный объект
Недолгий период развития от яйца до имаго, уникальное разнообразие мутационных рас с характерно отчетливым проявлением, всего 4 хромосомы и ряд других особенностей делают дрозофилу незаменимой для проработки закономерностей наследственности на практике. Наши ученые – генетики, такие как И. А. Рапопорт, Н. П. Дубинин, их зарубежные коллеги – Т. Г. Морган, Г. Д. Миллер, Р. Эксел проводили свои эксперименты именно на плодовой мушке.
Исследования продолжаются и по сей день: считается, что дальнейшее изучение белков, ответственных за восприятие вкуса и запаха насекомыми, поможет разработать препарат, способный сделать сельскохозяйственные культуры невкусными для насекомых – вредителей.
Сложно передать словами удовольствие, которое испытываешь, рассматривая этих мушек в бинокулярный микроскоп с небольшим увеличением. Эти переливающиеся крылышки и загадочные глаза привели когда-то титанов науки к величайшим открытиям. Аромат эфира. Видимо, знаком не одному сотруднику генетической лаборатории, ведь он необходим для усыпления непоседливых мушек, а, следовательно, для нормальной работы.
Итак, не бойтесь взглянуть дрозофиле в глаза, ведь так много тайного сможет нас объяснить эта неприметная мушка.
Список литературы:
Модельные объекты в нейробиологии: позвоночные
Автор
Круг объектов исследования современной нейробиологии огромен, однако далеко не все из них стали модельными для решения фундаментальных и прикладных задач, стоящих перед этой областью естественных наук. Наиболее традиционными модельными объектами нейробиологии долгие годы были представители позвоночных животных и прежде всего млекопитающих (крысы, мыши, кролики, собаки и кошки). Это в значительной степени связано со сходством организации их нервной системы с нервной системой человека. Удобство содержания в лабораторных условиях, плодовитость, быстрый рост, более детальная изученность их организации в целом сделали особенно крыс и мышей классическими модельными объектами для всестороннего изучения нервной системы высших животных, их поведения, механизмов памяти, последствий воздействия на нервную систему и сенсорные органы повреждающих факторов, механизмов функциональной и посттравматической репарации нервной системы, ее онтогенеза. Для максимально гуманного использования подопытных животных в научно-исследовательских работах в ряде стран были разработаны специальные рекомендации по содержанию животных, постановке с ними экспериментов, обезболиванию и наркотизации. Соблюдение этих рекомендаций в значительной степени стало контролироваться.
Постепенно с получением новых знаний о нервной системе представителей других групп позвоночных круг модельных объектов нейробиологии был расширен за счет введения в него представителей птиц, амфибий, рептилий и рыб. Это позволило дополнительно на примере более детального изучения отдельных видов выявить основные закономерности и особенности эволюционных преобразований нервной системы и сенсорных структур в разных таксономических группах позвоночных (см. раздел Животные).
Способность животных воспринимать мельчайшие изменения в окружающей среде с помощью своих органов чувств всегда вызывала пристальное внимание нейробиологов. Поскольку на человеке невозможно проводить полноценное изучение структурно-функциональной организации сенсорных систем, механизмов кодирования и обработки сенсорной информации, а также связи сенсорной периферии с высшими отделами мозга и формированием ими моторных команд, то все это стали успешно исследовать на представителях позвоночных животных. Благодаря этим работам были получены выдающиеся результаты в изучении молекулярных механизмов зрительной, обонятельной и вкусовой рецепции, физиологии интероцептивного анализатора, механизмов внутриклеточной сигнализации, вестибулярных функций и мн. др. Полученные данные легли в основу разработки теории передачи и восприятия изображений, моделирования обонятельной и зрительной функций с целью создания технических датчиков и т.п.
Содержание



Рекомендуемая литература
Врановые птицы как модельный объект для изучения нервной системы
Автор
Редактор
Врановые в настоящее время широко используются в качестве объекта для изучения высшей нервной деятельности, основ мышления и эволюции центральной нервной системы. Это направление исследований было заложено в работах Л.В. Крушинского (на Кафедре высшей нервной деятельности МГУ, г. Москва), в которых он впервые показал на ряде представителе млекопитающих и птиц, что у животных существуют элементы рассудочной деятельности. Изучение высшей нервной деятельности птиц, чьи способности превосходят хищных млекопитающих и достигают уровня приматов, позволяет понять истоки возникновения в эволюции позвоночных животных и человека мышления и сознания (Зорина, Полетаева, 2002; Зорина, Обозова, 2011).
В пределах класса птиц были обнаружены сходные с млекопитающими градации способности к экстраполяции – от полного её отсутствия у голубей до высокого её развития у врановых птиц. Для изучения способности птиц к логическому мышлению, использовали различные тесты: эксперименты с непрозрачной ширмой и кормушками, тест Ревеша–Крушинского, тест на способность животных к сравнению множеств и т.п.). Во всех этих тестах врановые показали результаты, сравнимые с приматами.
Результаты всех этих тестов позволили сделать вывод о том, что представители группы врановых способны улавливать логическую структуру задач (Багоцкая и др., 2010). Кроме того, в их сообществах выявлена чрезвычайно сложная социальная структура и общественное поведение (Баккал, 1997; Зорина и др., 1999). Результаты исследований свидетельствуют о наличии у них целого ряда высших когнитивных функций, в том числе и об их способности к счету, употреблению и даже изготовлению орудий труда (Багоцкая и др., 2013; Зорина, Салимов, 1989; Зорина, Обозова, 2011; Зорина, Полетаева, 2002; Hunt, 1996; Hunt, Gray, 2003). Например, Новокаледонский ворон Corvus moneduloides известен чрезвычайной изобретательностью при добывании пищи. Из растительных материалов он делает крюкообразные или штыкообразные приспособления, с помощью которых достаёт из-под коры личинок. Изготовленные орудия ворон может забирать с собой, пользуясь ими в дальнейшем. При изучении поведения животных и их социальной жизни используют обычно телевизионные камеры.
Интересно, что способность к рассудочной деятельности у птиц обеспечивается головным мозгом, существенно отличающимся по организации от мозга млекопитающих. Наиболее значимые различия выявлены в строении больших полушарий птиц и млекопитающих (Обухов, 1999, 2005). Все это позволяет использовать представителей врановых в качестве модельных объектов для изучения структурных основ, особенностей и общих закономерностей становления и развития в ходе эволюции животных и человека высшей нервной деятельности.
Рекомендуемая литература
Рыбы как модельный объект в нейробиологии, биологии развития и биотехнологии
Многие представители рыб являются незаменимыми модельными объектами для нейробиологических исследований. Рыбы являются крупнейшим классом позвоночных животных и той группой, которая дала начало наземным позвоночным животным, поэтому изучение их нервной системы имеет важное значение для понимания закономерностей эволюции нервной системы у позвоночных в целом. Кроме того, широкая адаптивная радиация среди современных рыб дает возможность исследовать влияние факторов среды на формирование вариантов в структуре разных отделов мозга, оценить уровень воздействия этих факторов на структурно-функциональные параметры нервных центров, их связь с развитием органов чувств. В связи с этими задачами для изучения в качестве модельных объектов отбираются виды рыб из различных филогенетических и экологических групп. В настоящее время для таких исследований используют представителей практически всех основных подклассов и отрядов рыб. Из Хрящевых рыб это в первую очередь акулы – катран Squalus acanthias L. и кошачья акула Scyliorhinus canicula L. Из Костных рыб – данио Danio rerio Hamilton 1822 и сазан Cyprinus carpio Linnaeus, 1758. Широко используют и ряд видов осетровых и лососевых рыб.
Интерес к исследованию центральной нервной системы рыб обусловлен также тем, что они являются удобным объектом для экспериментального исследования процессов постнатального нейрогенеза в нервной системе позвоночных животных. Показано, что в ряде отделов их головного мозга во взрослом состоянии присутствуют пролиферативные зоны, где обнаружены клетки так называемой радиальной глии. Эти клетки в настоящее время рассматриваются в качестве нейрональных предшественников не только в мозге рыб, но и в мозге млекопитающих животных и человека (см. подробнее раздел Постэмбриональное развитие позвоночных).
Использование рыб позволило разработать методические подходы для инструментального анализа структурных изменений в нервной ткани при нормальном развитии центральной нервной системы и при травме. Разработка новых технологий на основе изучения механизмов репарации мозга у различных животных и, в частности, рыб является инновационным подходом, благодаря которому могут быть по-новому оценены факторы, ограничивающие регенерационные возможности мозга млекопитающих и человека, а также найдены новые подходы к лечению повреждений мозга высших животных и человека.
Рыбы успешно используются также и в качестве модельных объектов для изучения воздействия повреждающих факторов окружающей среды на развитие мозга и сенсорные структуры. В первую очередь это относится к исследованиям последствий загрязнения природных водоемов продуктами нефтедобычи и нефтепеработки (мазут, бензин, дизельное топливо). В условиях интенсивного развития нефте- и газодобычи на морских шельфах, а также увеличения загрязнения внутренних водоемов продуктами нефтепереработки эта проблема становиться все более актуальной. Особенно это важно для сохранения видового разнообразия рыб, естественного и искусственного воспроизводства популяций ценных пород рыб (осетровых, лососевых). Полученные в таких исследованиях экспериментальные данные могут быть использованы в качестве тест-показателей степени влияния загрязняющих факторов среды на развитие организмов и стать основой для разработки ГОСТов на предельно-допустимые концентрации (ПДК) загрязняющих веществ для рыбоводческих целей.

Поперечный разрез через туловищный мозг стрекозы (А) и человека (В) со схемой нейрональных отношений (по Заварзину, 1950, с небольшими изменениями).
Синим цветом изображены двигательные (эфферентные) нейроны; красным – чувствительные волокна; коричневым – ассоциативные нейроны, связанные с двигательной областью; оранжевые и зеленые – собственный ассоциативный аппарат данной зоны (местные нейроны, фиолетовые – вторичные пути чувствительной области; Fa-передняя щель, sp-задняя септа; Dw и Vw – дорсальные и вентральные корешки, остальные обозначения относятся к разным типам клеток и волокнам)



Рекомендуемая литература
Модельные объекты нейробиологии: беспозвоночные животные
Автор
Относительно простые нервные системы многих беспозвоночных животных также оказались весьма эффективными моделями для всесторонних нейробиологических исследований. Этому в значительной степени способствовал открытый в начале прошлого века академиком А.А. Заварзиным принцип структурно-функционального параллелизма развития тканей многоклеточных организмов. Этот принцип стал одним из основополагающих в эволюционной морфологии и физиологи. В его основу легли полученные Заварзиным неоспоримые факты, свидетельствующие о наличии конвергентного сходства в строении функционально аналогичных систем у животных, далеко отстоящих друг от друга в филогенезе. Было показано, что за анатомически несхожими на первый взгляд оптическими центрами насекомых, головоногих моллюсков и позвоночных животных, а также туловищным мозгом позвоночных (спинной мозг) и членистоногих (брюшная нервная цепочка) стоит глубокая функциональная аналогия, проявляющаяся в цитоархитектонике и основных клеточных типах этих нервных структур. Заварзин установил, что спинной мозг и брюшная нервная цепочка демонстрируют сходство в своей нейрональной организации независимо от принадлежности изучаемой формы к тому или иному филогенетическому ряду животных, а только в зависимости от одинаковой высоты их развития, проявляющейся в определенном уровне их функциональных возможностей, что подчеркивает неразрывность в ходе эволюции формы и функции как двух сторон единого биологического процесса. В дальнейшем теория параллелизма академика А.А. Заварзина нашла свое широкое подтверждение не только на разных уровнях организации нервной системы и других функциональных систем многоклеточных организмов, но и на других уровнях живой материи.

Схемы нейрональных отношений в оптических путях разных по уровню организации и систематическому положению животных: А – насекомых по Заварзину, В – птиц по Рамон-и-Кахалу и С – каракатицы по Рамон-и-Кахалу (из Заварзина, 1950).
Синим обозначены нейроны первого оптического ганглия, красным – второго, зеленым – третьего; I, II, III – первый, второй и третьи оптические ганглии, oz – светочувствительные клетки глаз насекомого, a – слой палочек и колбочек в сетчатке птиц, rz – светочувствительные (ретинальные) клетки в глазу каракатицы, остальные обозначения относятся к разным типам нервных клеток их отросткам и связям
Все это сразу привлекло к гастроподам пристальное внимание широкого круга нейробиологов и наметило два основных направления в изучении их нервной системы: 1 – изучение свойств и механизмов функционирования нервной клетки и ее отдельных элементов и 2 – исследование морфо-функциональной организации нейрональных ансамблей и их звеньев, участвующих в осуществлении рефлекторных актов и основных форм поведения животных. К одним из первых работ по изучению морфологических и физиологических свойств отдельных нервных клеток мозга на примере идентифицированных нейронов центральной нервной системы гастропод относятся исследования висцерального (абдоминального) ганглия морского заднежаберного моллюска аплизии (Arvanitaki, Chalazonitis, 1955; Hyghes, Tauc, 1961 и др.), гигантских нейронов мозга нескольких видов голожаберных (Сахаров, 1965; Dorsett, 1967 и др.) и легочных моллюсков (Kerkut, Walker, 1974). Первые работы по изучению клеточных основ поведения на гастроподах были также предприняты на заднежаберных моллюсках и были связаны с клеточным анализом реакции уплывания от опасности или иначе комплекса фиксированных локомоторных действий у тритонии (Willows, 1971), организации некоторых простых висцеральных рефлексов и защитно-оборонительного поведения у аплизии, проявляющегося, например, в отдергивании сифона и выпускании чернильной жидкости (Кэндел, 1980; Шепперд, 1987 и др.).
Многочисленные исследования, проведенные к настоящему времени на нервной системе отдельных видов брюхоногих моллюсков и все возрастающее привлечение ее к решению широкого круга нейробиологических проблем, подтвердили перспективность и уникальность моллюсков как модельных объектов для выяснения фундаментальных свойств нервной системы. Благодаря исследованиям, проводимым на гастроподах, достигнуты большие успехи в изучении синаптических структур, нейротрансмиттеров и нейромодуляторов и их роли в интегрирующей и координирующей функции нервной системы (Сахаров, 1974, 1990; Дьяконова, 2012 и др.). С помощью гастропод получены принципиально новые данные по клеточным основам обучения и памяти (Кэндел, 1984; Балабан, Захаров, 1992; Гуйнудинова, Чекмарев, 1996 и др.). Составлены схемы организации и выявлен ряд механизмов функционирования нейрональных систем, обеспечивающих локомоцию (Аршавский и др., 1987; Frost, Kater, 1996), пищевое (Hurwitz, Susswein, 1996) и защитно-оборонительное поведение (Максимова, Балабан, 1983; Захаров, 1992), висцерокардиальные и респираторные рефлексы (Журавлев и др., 1994; Moroz, Winlow, 1992), а также принимающих участие в ряде половых актов (Кэндел, 1980) и многое другое.
На представителях легочных наземных улиток и слизней, обладающих хорошим обонянием, успешно изучают организацию и работу хемосенсорных систем и связанных с поступающей от них информацией нервных центров мозга (Chase 1986, 2002; Gelperin, Tank, 1990; Зайцева, 2016). Было показано, что степень развития тентакулярных обонятельных органов и всей обонятельной системы моллюсков связаны не только с филогенетическим положением и общим уровнем организации животного, но в значительной степени со средой его обитания и образом жизни, т.е. со степенью сложности и спецификой решаемых ими задач. В ходе эволюции выход на сушу и усложнение в связи с этим окружающей среды дали мощный толчок к прогрессивному развитию не только периферических, но и центральных отделов обонятельной системы у моллюсков. Это привело к неоднократному и независимому появлению в разных по уровню организации филогенетических группах брюхоногих, ведущих наземный образ жизни, сходной организации обонятельной системы, имеющей целый ряд общих черт с организацией обонятельных систем членистоногих и позвоночных животных и в частности появление, как у насекомых и позвоночных, обонятельных гломерул (Зайцева, 1989, 2000а, 2016; Zaitseva, 1991).
Полученные в ходе всех этих многочисленных исследований данные еще раз подтвердили универсальность многих свойств как отдельных нервных элементов, так и всей нервной системы в целом. Это отчетливо прослеживается, например в таких далеко отстоящих в филогенезе группах животных, которыми являются позвоночные, насекомые и брюхоногие и головоногие моллюски.

--> Соревнования по полевой ботанике "ВЕСЕННЯЯ ФЛОРА" пройдут в мае-июне 2020 года в онлайн-формате (определение растений по фотографиям). К участию в соревновании приглашаются школьники и взрослые любители природы, проживающие в средней полосе Европейской части России. Подробнее >>>
| [ sp ] : ml об : |
Наши авторские методические материалы по водной экологии и гидробиологии:
В нашем Интернет-магазине по некоммерческим ценам (по себестоимости производства)
можно приобрести следующие методические материалы по водной и околоводной флоре и фауне России:
Прудовик ушковый - Lymnaea auricularia
Прудовик ушковый относится к семейству Прудовиков (Lymnaeidae), которые обладают хорошо развитой, спирально закрученной вправо раковиной из отряда Лёгочных улиток (Pulmonata) класса Брюхоногих моллюсков (Gastropoda).
Синонимы. Прудовик ушковый - Lymnaea auricularia (Linnaeus, 1758), = Radix auricularia Linnaeus, 1758; = Helix auricularia Linnaeus, 1758, = Turbo patulus Da Costa, 1778, = Limnaeus auricularius Stein, 1850, = Radix tumida Ehrmann, 1933. По Я.И. Старобогатову - это 2 вида подрода Radix.
Внешний вид. Прудовики обладают хорошо развитой спирально закрученной вправо (см. как определить закрученность) раковиной (фото 1 и рис. 1). Из раковины высовывается передняя часть тела с широкой, поперечно обрезанной головой, явственно отделенной от тела шейным перехватом. На голове находятся подвижные щупальца - плоские, треугольной формы, короткие и широкие. У основания щупаалец сидят глаза. С брюшной стороны головы в передней её части помещается ротовое отверстие. Поверхность тела прудовика, соприкасающаяся с водой, покрыта ресничками.
Из раковины также высовывается нога - большой мускулистый орган движения, занимающий всю брюшную часть его тела. Нога у прудовиков довольно длинная и массивная. Если потревожить прудовика, например взять его в руки, то нога сократится и закроет устье раковины. В таком положении хорошо можно рассмотреть подошву ноги, уплощенную по карям и имеющую удлиненно-овальную форму. Клетки кожного эпителия на подошве обильно выделяют слизь, которая облегчает движение прудовика и способствует прилипанию подошвы к отвесной поверхности.
Раковина. Устье раковины ушкового прудовика по форме похоже на человеческое ухо, размер раковины меньше, чем у обыкновенного прудовика, ее высота - 2,5-3,5 см, ширина до 25 мм, высота устья до 30,3 мм, ширина устья до 21 мм, высота завитка до 5,4 мм.
Раковина серовато-желтого, желтого, желтовато-рогового, рогового или грязно-рогового цвета, обычно тонкостенная, хрупкая, полупрозрачная. Поверхность ее слегка блестящая, с тонкой неравномерной осевой исчерченностью и отчетливыми линиями приостановки роста, иногда с маллеатной скульптурой (уплощенные участки, похожие на следы от ударов молотка). Последний оборот очень большой (высота составляет 0,87 высоты раковины), неширокий, сильно вздутый.
Молодые раковины не имеют столь сильно раздутого последнего оборота, поэтому форма их не уховидная, а яйцеобразная. Шов, разделяющий обороты раковины, умеренно глубокий, скошенный. Оборотов до 4. Первые 3 оборота образуют очень короткий завиток, в результате он заостренный (узкий и острый), с вогнутыми контурами, выступает над устьем на 2-8 мм. Высота завитка меньше высоты устья, а у молодых особей равна приблизительно половине высоты устья. У живого моллюска раковина никогда не бывает прикрыта мантией, в отличие от похожей по внешнему виду раковины плащеноски, верхний край устья имеет горб или горизонтальный участок.
Тело. Тело, голова и нога ушкового прудовика зеленовато-серого, желтовато-оливкового или оливкового цвета с многочисленными мелкими беловатыми, желтыми или темными крапинками, более или менее равномерно разбросанными по их поверхности. Мантия серовато-, светло- или желтовато-оливковая, однотонная, чаще - пятнистая (со светлыми или темными пятнами). Щупальца желтоватые или беловатые.
Внутреннее строение. Глотка прудовика представляет мускулистый мешок, который переходит в пищевод, затем в зоб и желудок. Желудок состоит из двух отделов - двулопастного мускулистого отдела и удлиненного пилорического. Мускулистый желудок отличается грубостью строения и способствует размельчению захваченной пищи. В пилорическом желудке и в выходящей из него кишке пища переваривается. Анальное отверстие открывается в устье раковины.
Мышечных гребней и адаптивной жабры в легочной полости нет. Имеется короткий сифон, сформированный за счет наружного края мантии.
Строение остальных внутренних органов прудовиков в целом типично для моллюсков и подробно изложено в описании класса Брюхоногих (Gastropoda) и отряда Лёгочных улиток (Pulmonata).
Движение. Движения прудовиков бывают трех типов - ползание (скольжение) по поверхностям с помощью ноги, всплытие и погружение за счет лёгочной полости и скольжение снизу по поверхностной плёнке воды.
Ползание прудовика по подводным поверхностям вызывается мускульными сокращениями ноги, волнообразно и равномерно пробегающими по ее подошве. Движения эти обладают тонкой приспособляемостью, которая позволяет моллюску двигаться по тоненьким веточкам и листьям водных растений.
Всплытие и погружение осуществляется за счет наполнения и опустошения лёгочной полости. Дело в том, что удельный вес прудовика, содержащего воздух в лёгочной полости, меньше удельного веса воды и имея запас воздуха моллюск автоматически поднимается к поверхности воды, сложно изгибаясь при этом и далеко выставив ногу. Для погружения моллюск может произвольно сжимать свою дыхательную полость - тогда он становится тяжелее и опускается на дно. При расширении полости улитка по вертикальной линии всплывает на поверхность. Для экстренного погружения (например, в случае опасности), прудовик выталкивает находящийся в лёгочной полости воздух и резко падает на дно. После этого моллюск уже не в состоянии подняться на поверхность иначе, как переползая по подводным поверхностям.
Скольжение по нижней поверхности воды осуществляется после всплытия прудовика на поверхность. Прудовик касается подошвой ноги плёнки поверхностного натяжения, затем обильно выделяет слизь, расправляет ногу, несколько выгибая подошву внутрь в виде лодочки и, сокращая мускулатуру подошвы, скользит по плёнке поверхностного натяжения, покрытой тонким слоем слизи. В ясный солнечный день можно видеть, как слизистая пленка, отливая разными цветами, блестит на солнце. Эта слизь может быть также обнаружена, если провести какой-либо палочкой позади ползущей улитки.
Дыхание. Как и другие лёгочные улитки, прудовики лишены первичных жабр и дышат атмосферным воздухом с помощью лёгкого - специализированного участка мантийной полости, к которому прилегает густая сеть кровеносных сосудов. Мантия прудовика представляет собой складку кожи, края которой, образуя полость, срастаются со стенкой тела улитки, за исключением дыхательного отверстия, расположенного сбоку тела, близ края раковины, и снабженного кольцевой мускулатурой, открывающей и закрывающей вход в мантийную полость. Для того, чтобы обновить воздух в лёгочной полости, прудовики периодически поднимаются к поверхности воды. Поднявшись на поверхность, прудовик открывает свое дыхательное отверстие и воздух втягивается в мантийную полость. В это время можно услышать характерный хлюпающий звук - "голос моллюска".
При опускании в воду воздух в полости мантии содержит количество кислорода, близкое к атмосферному, но под водой это количество быстро потребляется. Поэтому прудовик вынужден постоянно подниматься на поверхность за очередной порцией воздуха. Частота подъёмов для дыхания зависит от температуры воды. В хорошо прогретой воде при температуре 18-20° прудовики поднимаются на поверхность по 7-9 раз в час. Так дышит прудовик летом.
По мере того, как температура воды понижается, они начинают всплывать на поверхность все реже и осенью еще задолго до замерзания водоема при температуре 6°-8° C, в связи с общим падением активности, перестают вовсе подниматься на поверхность. Пока фотосинтез водных растний продолжается, прудовики потребляют для дыхания пузырьки кислорода на растениях, а затем перестают наполнять воздухом полость мантии. При этом она или спадается или наполняется водой и начинает функционировать как жабры - т.е. растворенный в воде кислород переходит в кровь моллюска напрямую через складки мантии. Такой способ дыхания является парадоксальным и редким в природе явлением, когда один и тот же орган попеременно функционирует то как жабры, то как легкое.
Такой способ дыхания наблюдается чаще всего у мелких прудовиков, которые обитают в чистой, богатой кислородом воде на достаточно больших глубинах, откуда им трудно подниматься к поверхности. У них лёгкое заполняется водой и фунционирует как жабры. Моллюски, обитающие в таких условиях, как правило, мельче представителей того же вида, населяющих мелководья.
За счет преимущественно атмосферного воздуха дышат, в основном, те прудовики, которые живут в мелких, заросших растительностью водоёмах - в лужах, в прудах, в области зарослей озер и рек. Ползая среди растений поблизости от поверхности воды, прудовики легко поднимаются к поверхности для дыхания атмосферным воздухом.
Помимо воздушного и водного дыхания, протекающих в полости легкого, прудовик живет также и за счет кожного дыхания, которое выполняется всей поверхностью тела, омываемого водой. При этом большое значение имеют реснички кожного покрова прудовика, непрерывное движение которых способствует току воды, омывающей поверхность тела моллюска. Особенно большое значение кожный тип дыхания имеет для прудовиков, живущих в области прибойного побережья озер и водохранилищ, где они сидят плотно прижавшись к поверхности камней или к колеблемым ветром стеблям тростника с помощью своей сильной ноги. Прудовики в прибойной зоне никогда не поднимаются на поверхность воды для дыхания. Это было бы для них губительно, так как прибой разбил бы о камни их нежные раковины или вовсе выбросил бы их из воды, что и без того нередко наблюдается после бури в малых озерах, когда множество прудовиков вместе с водными растениями оказывается вынесенными на берег.
Питание. Прудовики всеядны, но в природе предпочитают растительную пищу. Медленно ползая, они соскабливают налеты водорослей с различных погруженных в воду предметов, например с поверхности стеблей и листьев высших водных растений. Если водорослей становится мало, то они потребляют и живые растения - листья и стебли водных растений, выбирая наиболее нежные из них, а также растительный детрит.
Для соскабливания пищи прудовики пользуются зубчатой тёркой - роговой пластинкой, расположенной в глотке и усаженной рядами зубчиков (см. рис. 2: 3 в описании семейства Прудовиков). Способ работы терки легко наблюдать в аквариуме, когда прудовик ползёт по стеклу и время от времени высовывает тёрку изо рта наружу и проводит ею по поверхности стекла, чтобы соскоблить слой развившихся на нем зелёных водорослей.
Прудовики питаются иногда и животной пищей - поедают трупы головастиков, тритонов, рыб и моллюсков, скобля их с поверхности, а также заглатывая целиком мелких беспозвоночных животных (гидр, простейших) и рыбью икру.
Размножение и развитие. Прудовики, подобно прочим Лёгочным брюхоногим, гермафродиты. Яйца и сперматозоиды развиваются в одном организме, в различных участках одной и той же железы, но по выходе из нее пути половых протоков разделяются, и мужское и женское половые отверстия около устья раковины открываются раздельно.
Из мужской половой поры во время копуляции выступает мускулистый копулятивный орган, женская половая пора ведет в обширный семеприемник. У прудовиков наблюдается спаривание, причем одна особь играет роль самки, а другая самца, или же оба моллюска взаимно оплодотворяют друг друга. Иногда образуются цепи копулирующих прудовиков, причем крайние особи выполняют роль самки или самца, а срединные - и того и другого.
Откладка яиц продолжается в течение всего теплого времени года, начиная с ранней весны, а в аквариуме и зимой. Яйца прудовиков в отложенном состоянии связаны общей слизистой оболочкой. Кладка имеет вид прозрачной студенистой колбаски с округлыми концами, которую моллюски откладывают на водные растения или другие предметы.
Дней через 20 из икринок выходят крошечные улиточки, уже снабженные раковиной, которые довольно быстро растут, питаясь растительной пищей.
Прудовики могут размножаться, далеко не достигнув своего предельного возраста и размера. Чаще всего они становятся половозрелыми уже в конце первого года своей жизни, когда он вырастает лишь в половину своей нормальной величины. Прудовики могут размножаться и будучи изолированными от других особей, так что копуляция не представляет у них акта, необходимого для продолжения жизни, размножение вполне может происходить путём самооплодотворения.
Образ жизни и местообитания. В разгар лета прудовики держатся близ поверхности водоёма, а иногда и на самой поверхности воды. Для их лова нет даже надобности пользоваться сачком, они легко могут быть сняты с подводных предметов рукой.
При высыхании водоемов, заселенных прудовиками, таких как мелкие озерца, канавы и лужи, далеко не все моллюски погибают. При наступлении неблагоприятных условий моллюски выделяют плотную пленку, замыкающую отверстие раковины. Некоторые могут переносить пребывание вне воды довольно долго - от двух недель до месяца.
Зимой, при замерзании водоемов моллюски не погибают, а вмерзают в лед и оживают при оттаивании.
Ушковый прудовик населяет прибрежье самых различных водоемов - от прудов и озер до рек и ручьев. Держится на растениях, камнях, корягах, иле, встречается довольно часто.
Хозяйственное значение. Большинство прудовиков являются животными либо безразличными для человека, либо даже полезными для него, так как служат пищей для рыб.
Другие особенности строения, движения, питания, дыхания, размножения и развития, образа жизни, мест обитания и распространения, а также значения в жизни человека, характерные для всех прудовиков, читайте в описании семейства Прудовики - Lymnaeidae.
Наши авторские методические материалы по зоологии беспозвоночных:
В нашем Интернет-магазине по некоммерческим ценам (по себестоимости производства)
можно приобрести следующие методические материалы по беспозвоночным животным России:
В разделе Методические материалы Вы также можете познакомиться с описаниями разработанных экологическим центром "Экосистема" печатных определителей растений средней полосы, карманных определителей объектов природы средней полосы, определительных таблиц "Грибы, растения и животные России", компьютерных (электронных) определителей природных объектов, полевых определителей для смартфонов и планшетов, методических пособий по организации проектной деятельности школьников и полевых экологических исследований (включая книгу для педагогов "Как организовать полевой экологический практикум"), а также учебно-методических фильмов по организации проектной исследовательской деятельности школьников в природе. Приобрести все эти материалы можно в нашем некоммерческом Интернет-магазине. Там же можно приобрести mp3-диски Голоса птиц средней полосы России и Голоса птиц России, ч.1: Европейская часть, Урал, Сибирь.
Читайте также: